構造分子薬理学分野ではこんなテーマをやっています
- 生命活動を理解する上で,その構成成分である蛋白質の「かたち」を調べることは大きな助けになります.蛋白質の重要な性質の一つは「分子認識」をすることです。蛋白質の水溶液中での構造や、他の分子との相互作用の様子を解析するのに適した核磁気共鳴法(NMR)を主に使います。それにX線結晶解析法やクライオ電子顕微鏡などを組合わせて研究を進めます.
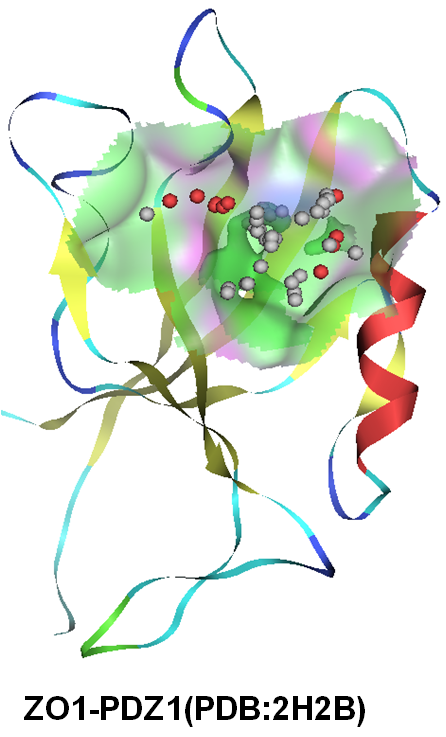
- 上皮タイトジャンクション裏打ちタンパク質を標的とした構造に基づいた創薬 (SGDD)
廣明グループ最重要テーマです。マ ルチPDZドメインタンパク質には細胞接着装置(上皮タイトジャンクション)の裏打ち(足場)タンパク質や、細胞骨格・細胞極性の制御因子など、多くの重 要な因子が多く含まれています。我々のグループでは既にいくつかのPDZドメインの構造決定を完了し、またそれに結合する新規の低分子化合物、たとえばタイトジャンクションを増強する化合物とタイトジャンクションを減弱しバリアを開放する化合物(医薬品吸収補助剤候補物質)などを見出しています。併せて、類似の化学構造を持ち、SARS-CoV2を阻害する化合物のSGDD研究も進めています。(廣明・天野・安藤・岡田)。
化学的架橋により糖部コンフォメーションを固定した非天然オリゴヌクレオチドは、生体内で通常の核酸よりもすぐれた性質を示します。その物性・熱力学的安定 性を研究しながら、アンチセンス医薬品・アプタマー医薬品などの次世代の核酸医薬品を設計し、実用化を目指します(兒玉・伊藤・野中・杉浦・山内)。
天然変性タンパク質(IDP)は高等真核生物のゲノム中に多い(それも核内タンパク質に特に多い)生理条件下で単独では立体構造をとらないタンパク質の配列です。廣明グループでは、天然変性タンパク質に、他の酵素に対する凍結保護効果・凍結乾燥保護効果があることを発見し、2015年1月にはJSTの支援の下、国際特許を出願しました。現在はIDPが関連する感染症関連タンパク質を標的とした創薬研究を、AMED-CRESTプロジェクトに参画し、新潟大学・松本教授の指導の下進めています(廣明・合田・天野・浅井・胡)。
p53はおよそ半数のがんで変異が見つかる、正常細胞の分裂抑制や異常化した細胞のアポトーシスを誘導する「がん抑制因子」の一つです。p53遺伝子の不思議の一つは、細胞内に二つある遺伝子の片側に変異が入っても、正常な残り一方由来の遺伝子産物も巻き込んで、p53タンパク質が不活性化してしまうdominant negative現象が知られていることです。私たちはそのメカニズムに、p53の形成する多様な凝集体が関わっていると考え、p53多量体の形成や検出のための物理化学的研究を進めています。(日比野・吉田)
RNaseH1は、DNA:RNAハイブリッド鎖という、特殊な立体構造をした核酸を選択的に切断する酵素です。ヒトRNaseH1は、その重要性に比して、その研究が進んでいません。しかし、核酸医薬品の一つであるアンチセンス核酸が、ヒト細胞内で作用するときに、RNaseH1の活性を利用しているという報告があるため、その機能解析は核酸医薬開発に欠かせません。また、HIVウイルスの逆転写酵素中のRNaseHドメインなどが新規の創薬標的であることから、その重要性は増しています(廣明・合田)。
リン酸化,ユビキチン化、SUMO化や蛋白質分解などの蛋白質翻訳後修復に関わる因子とその後の運命決定に係る分子群の構造解析と相互作用解析の研究を行っています。このうちタイトジャンクションの主要成分クローディンを特異的にユビキチン化する酵素LNX1については、その阻害剤を開発し、バリア強化剤として実用化研究も進めています(天野・廣明)。
京都大学医学研究科・TAOヘルスライフファーマ(株)と協力して、Abアミロイドペプチドのオリゴマー仮説に関連し、高毒性を示すAb凝集体(アミロスフェロイド:ASPD)のNMRによる構造解析、阻害剤設計、抗体との相互作用解析などを行います。またアミロイド阻害薬のNMRスクリーニング研究も行っています(廣明・合田)。
膜骨格・脂質認識に関連する哺乳動物由来の蛋白質の構造生物学を進めます。特に赤血球膜裏打ちタンパク質ストマチンの形成する新規の線維構造や、細胞膜やオルガネラ膜の制御や構造変換に関与する分子群やAAA-ATPase、微小管切断酵素カタニンとその調節因子に関与する蛋白質の解析を行います(廣明)。
(2025年4月2日更新)